Nuevas aproximaciones a los mecanismos tumorales
Los mecanismos generadores de inestabilidad cromosómica juegan un papel clave en el desarrollo tumoral ya que permiten una continua remodelación del genoma. La inestabilidad cromosómica se puede definir como la continua aparición de ganancias y/o pérdidas de fragmentos cromosómicos y/o cromosomas enteros. Los modelos actuales de desarrollo tumoral postulan que la inestabilidad cromosómica se produciría al inicio del proceso neoplásico (tumoral) y que el continuo remodelado del genoma permitiría la aparición de mutaciones que otorgarían a las células las características necesarias para convertirse en malignas.
Una manera sencilla y rápida que permite determinar la presencia de inestabilidad cromosómica es el análisis de anomalías en la morfología de los núcleos de las células durante la interfase. Diferentes tipos de morfologías nucleares anómalas como los micronúcleos (MNS), las protuberancias nucleares (Bs) y los puentes nucleoplasmáticos (NPB) pueden manifestarse durante la interfase celular (imagen de más abajo) y, dependiendo de su fuente de origen, puede prevalecer más una que otra.
La pérdida de protección de los extremos cromosómicos o telómeros, estado conocido como disfunción telomérica, parece constituir uno de los mecanismos por los cuales las células son capaces de generar la inestabilidad cromosómica responsable de iniciar la carcinogénesis en modelos de ratón. En humanos también se han observado evidencias de que la disfunción telomérica puede tener un papel relevante como instigadora de la carcinogénesis. En este sentido, lesiones preneoplásicas y de estadios neoplásicos incipientes a menudo presentan cromosomas con telómeros excesivamente cortos y posiblemente disfuncionales. Además, varios estudios han asociado longitudes teloméricas críticamente cortas con un mal pronóstico tumoral. Así pues, es de relevancia caracterizar cuáles son los marcadores biológicos que indican la presencia de disfunción telomérica.
A tal fin, el trabajo que se presenta ha identificado y evaluado las morfologías nucleares anómalas en un ambiente con disfunción telomérica. El estudio se ha centrado en células epiteliales primarias humanas (HMECs) derivadas de reducciones estéticas mamarias, las cuales presentan niveles crecientes de disfunción telomérica a medida que se cultivan "in vitro". Previo al análisis y para preservar todas las anomalías morfológicas nucleares, las células fueron tratadas con citocalasina-B (CYT-B), una toxina que bloquea la división citoplasmática sin interferir con la división nuclear, estadios 1A-1D de la figura siguiente:
Bloqueo artificial de la citocinesis con citocalasina-B (CYT-B) y formación de anomalías en la morfología de los núcleos en células binucleada resultantes en la siguiente interfase. A) Célula binucleada sin morfología nuclear anómala. B) Célula binucleada con un puente nucleoplasmático (NPB). Esta anomalía mantiene los dos núcleos principales unidos por una tira fina de cromatina. C) Las protuberancias nucleares (B) son protrusiones que emergen del núcleo principal. D) Los micronúcleos (MN) son fragmentos de ADN o cromosomas enteros que han quedado físicamente excluidos de los núcleos principales. Mediante técnicas de hibridación in situ se marcaron concretamente los cromosomas 1-en rojo- y 22 -en verde- que presentaban un elevado grado de disfunción telomérica. Las imágenes B', C' y D' son ejemplos de cómo el cromosoma 1 forma parte de las diferentes anomalías morfológicas nucleares.
Las células binucleadas que resultaron exhibieron tres tipos diferentes de anomalías en la morfología de sus núcleos: Bs, NPBs y MNS, sin embargo, sólo los NPBs y Bs incrementaron de manera significativa paralelamente con la disfunción telomérica.
Para establecer de forma fehaciente una relación entre la presencia de anomalías en la morfología nuclear y la disfunción telomérica aplicaron técnicas de hibridación in situ, las cuales permiten identificar la posición de un cromosoma concreto en el núcleo celular. Las hibridaciones demostraron que los cromosomas con disfunción telomérica se encontraban más frecuentemente implicados en las morfologías del núcleo anómalas que los cromosomas con una longitud telomérica no disfuncional (Estadios 1B'-1D' de la anterior figura). Estos resultados claramente demostraban que la disfunción telomérica es una fuente de anomalías morfológicas nucleares, al tiempo que indicaban un posible origen común para todas ellas.
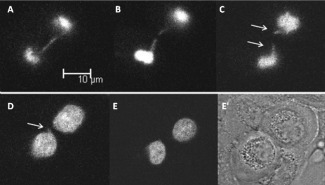
Para investigarlo, se hizo el seguimiento a tiempo real de mitosis en HMECs que contenían su ADN marcado con moléculas fluorescentes (GFP-H2B-HMECs). En estas células los cromosomas con telómeros disfuncionales a menudo se fusionan entre ellos originando cromosomas dicéntricos, los cuales originarán un puente de cromatina durante la anafase si las cromátidas hermanas son a la vez estiradas en direcciones opuestas hacia los polos de la célula, (figura de cabecera). El análisis del destino de estos puentes a partir de seguimientos de las células en tiempo real permitió determinar que la tensión ejercida sobre este puente es capaz de romperlo por un punto originando protuberancias nucleares y, de forma minoritaria, por dos puntos originando un micronúcleo.
En conclusión, nuestros resultados indican que la inestabilidad cromosómica resultante de disfunción telomérica se encuentra reflejada principalmente en la aparición de puentes nucleoplasmáticos y protuberancias nucleares, y minoritariamente en la aparición de micronúcleos. Y no sólo eso, sino que todo subraya la importancia del uso de estas anomalías como biomarcadores para determinar de manera fehaciente la presencia de disfunción telomérica en las células.
Referencias
"Telomere dysfunction and chromosome structure modulate the contribution of individual chromosomes in abnormal nuclear morphologies". Pampalona J, Soler D, Genescà A, Tusell L. Mutat Res. 2010 Jan 5;683(1-2):16-22. Epub . PubMed PMID: 19822157.